
2. El oído humano
El oído (figura 1) está formado por tres secciones: el oído externo, el oído medio y el oído interno, que pasaremos a describir desde los puntos de vista anatómico y funcional.
Figura 1. Corte transversal del oído derecho, en el cual se muestran las partes anatómicas más representativas del aparato auditivo.
Consta del pabellón u oreja, y el canal auditivo externo.
El pabellón recoge las ondas sonoras y las conduce hacia el canal auditivo mediante reflexiones y difracciones. Si bien es direccional, debido a sus irregularidades es menos direccional que la oreja de otros animales, como el gato o el perro, que además poseen control muscular voluntario de su orientación.
El canal auditivo, que mide unos 25 mm, conduce el sonido al tímpano. La parte más externa está recubierta por pilosidad y por glándulas sebáceas que segregan cerumen. Ejercen una acción higiénica, al fijar y arrastrar lentamente hacia el exterior las partículas de polvo que de otra forma se depositarían en el tímpano. Una segunda función es proteger al oído de ruidos muy intensos y prolongados, ya que la secreción aumenta en presencia de tales ruidos, cerrando parcialmente el conducto. Debido a la forma y las dimensiones físicas el oído externo posee una resonancia cuya frecuencia está en las proximidades de los 3000 Hz. Esta resonancia incide en la respuesta del oído, que luego estudiaremos.
Está ubicado en la caja timpánica, y lo integran el tímpano, los huesecillos u osículos, y la trompa de Eustaquio.
El tímpano es una membrana elástica, semitransparente y algo cónica, que comunica el canal auditivo externo con la caja timpánica. Es visible desde el exterior por medio del otoscopio (instrumento óptico que permite iluminar la zona a observar y está dotado a su vez de una lente de aumento). El tímpano recibe las vibraciones del aire y las comunica a los huesecillos. A causa de ruidos muy intensos (por ejemplo una potente explosión cerca del oído) o por determinadas infecciones, esta membrana puede perforarse, lo cual no es irreversible, ya que se cicatriza.
Los huesecillos son una cadena de tres pequeños huesos: el martillo, el yunque y el estribo (figura 2) que comunican al oído interno las vibraciones sonoras que capta el tímpano. Están sostenidos en su lugar por una serie de pequeños ligamentos y músculos. La finalidad de esta cadena es convertir vibraciones de gran amplitud y poca presión, como las hay en el tímpano, en vibraciones de pequeña amplitud y mayor presión, requeridas en el líquido que llena el oído interno. Esta función es asimilable, por consiguiente, a una palanca mecánica y se ilustra en la figura 2. A causa del efecto palanca las vibraciones del estribo son de menor amplitud pero mayor fuerza. La ganancia mecánica de esta palanca es de 1,3, lo que significa que la fuerza que el estribo ejerce sobre la ventana oval es 1,3 veces mayor que ejerce el tímpano sobre el martillo. A este efecto de palanca se agrega la gran diferencia de áreas entre el tímpano (0,6 cm2) y la ventana oval (0,04 cm2), lo cual implica que la relación entre las presiones en el tímpano y en la ventana oval es del orden de
![]()
Esta diferencia de presiones es necesaria ya que en el tímpano existe una impedancia acústica mucho menor que en el oído interno, ya que éste último contiene agua. El conjunto actúa, por consiguiente, como un ingenioso adaptador de impedancias acústicas.

Figura 2. Izquierda, los tres huesecillos del oído medio. Derecha, su mecánica.
Los músculos, además de la función de sostén de la cadena osicular, sirven de protección del oído interno frente a sonidos intensos. Cuando penetra en el oído un ruido muy intenso, se produce la contracción refleja de estos músculos, rigidizando la cadena, que pierde entonces su eficiencia mecánica, y la energía es disipada antes de alcanzar el oído interno. Esta protección sólo es efectiva, sin embargo, para sonidos de más de 500 ms de duración. En caso de lesión o esclerosamiento (endurecimiento) de la cadena osicular sobreviene un tipo de hipoacusia (sordera) caracterizado por una audición aérea disminuida y ósea normal. En la actualidad es posible reemplazar quirúrgicamente algunas partes por prótesis plásticas.
La trompa de Eustaquio es un pequeño conducto que comunica la caja timpánica con la laringe. Su función es la de igualar la presión del oído medio con la presión atmosférica. Normalmente, permanece cerrada, abriéndose en forma refleja durante la acción de tragar o de bostezar. Si permaneciera siempre abierta, el tímpano vibraría con una amplitud muy pequeña, ya que el movimiento del tímpano es el resultado de una diferencia de presión a uno y otro lado de éste. Por consiguiente, para vibrar en concordancia con las variaciones de presión sonora es preciso que la presión dentro de la caja timpánica se mantenga constante. Si, en cambio, la trompa de Eustaquio estuviera cerrada permanentemente (o no existiera), al variar la presión atmosférica se produciría una diferencia de presiones medias que curvaría el tímpano hacia afuera o hacia adentro, como se indica en la figura 3, reduciendo notablemente la respuesta auditiva, particularmente para los sonidos agudos (altas frecuencias). Esto es lo que sucede en los cambios bruscos de presión que se dan, por ejemplo, al ascender una montaña o al sumergirse varios metros debajo del agua, y el efecto es una sensación de embotamiento. La Naturaleza ha provisto la trompa de Eustaquio, que se abre al tragar, acción que se realiza periódicamente, ya sea en forma voluntaria o involuntaria, debido a la secreción permanente de saliva. La apertura de la trompa de Eustaquio provoca un ruido similar a un pequeño crujido, que el cerebro generalmente no tiene en cuenta, salvo que se preste especialmente atención.
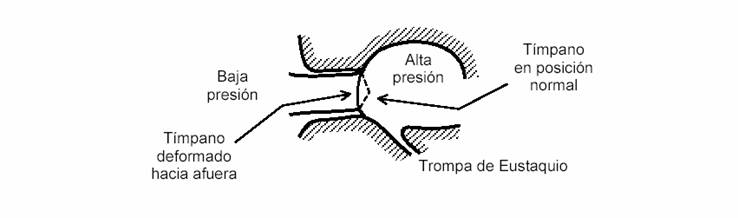
Figura 3. Deformación de la membrana timpánica ante una disminución relativamente rápida de la presión atmosférica.
Resumiendo, el objetivo de la trompa de eustaquio es independizar la audición de la presión atmosférica.
Está constituido por el laberinto, cavidad ósea que contiene a los canales semicirculares, el vestíbulo, y el caracol.

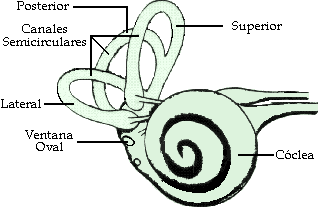
Figura 4. Aspecto idealizado del oído interno. En realidad el caracol y el laberinto son una cavidad en el hueso temporal
Los canales semicirculares son el órgano sensor del sistema de equilibrio. Son tres pequeños conductos curvados en semicírculo, con ejes aproximadamente en cuadratura. Interiormente están recubiertos por terminaciones nerviosas y contienen líquido endolinfático. Al rotar la cabeza en alguna dirección, por inercia el líquido tiende a permanecer inmóvil. Se crea un movimiento relativo entre el líquido y los conductos que es detectado y comunicado al cerebro por las células nerviosas, lo cual permite desencadenar los mecanismos de control de la estabilidad. Al haber tres canales en cuadratura se detectan movimientos rotatorios en cualquier dirección.


Figura 5. Funcionamiento de los tubos semicirculares.
El vestíbulo comunica los canales semicirculares con el caracol, y al mismo tiempo comunica el caracol con la caja timpánica a través de dos orificios denominados ventana oval y ventana redonda (también llamada tímpano secundario), cubiertos por sendas membranas de unos 3 mm y 2 mm respectivamente (figura 4). El estribo, última pieza de la cadena osicular, se encuentra adherido a la ventana oval.
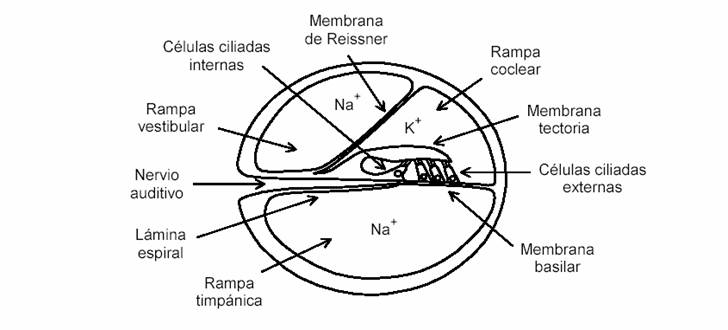
Figura 6. Corte transversal del conducto coclear en escala 25:1.
El caracol contiene el órgano principal de la audición: la cóclea, que es un tubo arrollado dos vueltas y media en espiral. Tal como se muestra en la figura 6, está dividida en tres secciones. La sección inferior, denominada rampa timpánica y la superior, conocida como rampa vestibular, contienen líquido perilinfático, rico en sodio (Na) y se conectan a través de un pequeño orificio, el helicotrema, ubicado hacia el vértice (ápex) del caracol. La cavidad central es la partición coclear o rampa coclear y contiene líquido endolinfático, rico en potasio (K).
La rampa vestibular se comunica con el oído medio a través de la ventana oval, y la rampa timpánica lo hace a través de la ventana redonda. La partición coclear contiene la membrana basilar, una membrana elástica sobre la que se encuentra el órgano de Corti, una estructura que contiene las células ciliadas o pilosas (figura 6). Las células ciliadas se comportan como diminutos micrófonos, generando pulsos eléctricos (denominados potenciales de acción) de unos 90 mV como respuesta a la vibración. Estos pulsos son enviados al cerebro a través de una serie de células nerviosas (neuronas) reunidas en el nervio auditivo. El potencial de acción de una célula individual no es fácil de medir, pero es posible medir la suma de ellos, aplicando unos electrodos transtimpánicos entre las ventanas oval y redonda. Estas tensiones se denominan microfónicos cocleares, y fueron observadas por primera vez por Wever y Bray, en 1930 en gatos.
La membrana basilar mide alrededor de 35 mm de longitud y tiene unos 0,04 mm de ancho en su zona basal (la más próxima a la base del caracol) y unos 0,5 mm en la zona apical (próxima al vértice o ápex). Además, la zona más angosta es también más rígida, lo cual será importante para la capacidad discriminatoria de frecuencias del oído interno. En la figura 7 se muestran dos vistas de la membrana con la cóclea hipotéticamente estirada desde su forma helicoidal hasta una forma rectilínea.

Figura 7. Dos vistas de la cóclea hipotéticamente rectificada. Arriba, vista superior. Abajo, vista lateral.
Cuando llega una perturbación a la ventana oval, el líquido de la sección superior se encuentra inicialmente a mayor presión que el de la sección inferior, lo cual provoca una deformación de la membrana basilar que se propaga en forma de onda (denominada onda viajera) desde la región basal hasta la región apical, tendiendo a aumentar la amplitud conforme la rigidez de la membrana va disminuyendo. Cuando la perturbación es periódica, tal como sucede con una vibración sonora, la membrana comienza a vibrar con una envolvente (figura 8) cuyo máximo se produce en cierta posición que depende de la frecuencia del sonido, como se muestra en la figura 9.

Figura 8. Arriba, onda viajera en la membrana basilar en un instante dado. Abajo, posición de la onda en tres instantes de tiempo t1, t2 y t3. Las líneas de trazos indican el lugar geométrico de los picos de la onda conforme ésta va avanzando a lo largo de la membrana.

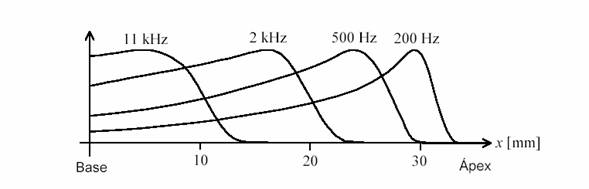
Figura 9. Envolvente espacial de las ondas viajeras sobre la membrana basilar para cuatro frecuencias diferentes.
Resulta, así, que existe una localización del pico de resonancia de la membrana basilar en función de la frecuencia, que se ha representado gráficamente en la figura 10. Esto confiere al oído interno una cualidad analítica que es de fundamental importancia en la discriminación tonal del sonido, especialmente para los sonidos de frecuencias superiores a los 1000 Hz. El descubrimiento de la mecánica de la membrana basilar se debe a Georg Békésy.
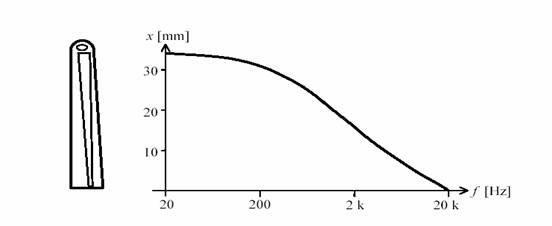
Figura 10. Ubicación de la resonancia a lo largo de la membrana basilar en función de la frecuencia
Como ya se anticipó, el movimiento de la membrana basilar ocasiona que las células ciliadas emitan un pulso eléctrico. El mecanismo para ello se ilustra en la figura 11. Debido a que las membranas basilar y tectoria tienen ejes diferentes, el movimiento relativo provoca un pandeo de los cilios que fuerza la apertura de unas diminutas compuertas iónicas. El intercambio iónico genera una diferencia de potencial electroquímico que se manifiesta como un pulso de unos 90 mV de amplitud o potencial de acción.

Figura 11. A la izquierda, una célula ciliada entre las membranas basilar y tectoria en estado de reposo. A la derecha, cuando se produce un movimiento de la membrana basilar a causa de una onda viajera, los cilios (pelos) de la célula ciliada experimentan un pandeo.
El potencial de acción generado por cada célula ciliada debe ser comunicado al cerebro. Ello se realiza a través de las neuronas. En primera aproximación podría imaginarse las neuronas como simples conductores eléctricos. Un análisis más detallado revela que son en realidad complejos sistemas con varias entradas y varias salidas, capaces de realizar operaciones de ponderación, de comparación y de generación de nuevos potenciales de acción. En la figura 12 se ilustra la estructura de una neurona típica, en la que se indican también los contactos con otras neuronas previas y ulteriores.
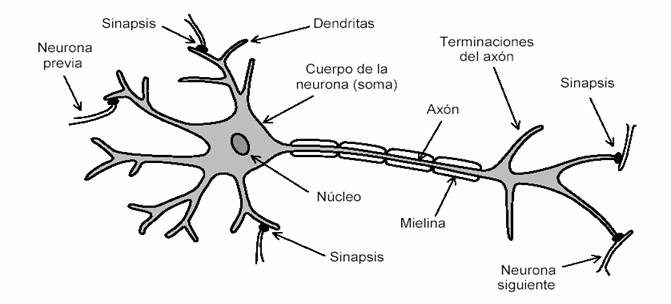
Figura 12. Una neurona típica, en la que se aprecian las dendritas que reciben potenciales de acción de las neuronas previas (o de las células sensorias, como las células ciliadas) a través de las sinapsis, el cuerpo o soma con su núcleo, el axón recubierto con mielina, y sus terminaciones que conectan con las dendritas de nuevas neuronas.
La neurona recibe potenciales de acción provenientes de neuronas anteriores o de células sensorias (como las células ciliadas) a través de las dendritas. Los puntos de contacto se denominan sinapsis. Las dendritas realizan una especie de suma ponderada de las señales recibidas, y si ésta excede cierto umbral, el propio cuerpo de la neurona descarga un nuevo potencial de acción de 90 mV y unos 2 ms de duración. Al terminar este pulso sobreviene un tiempo durante el cual no puede volver a emitir pulsos, denominado período refractario. El potencial de acción generado se propaga en forma de onda a lo largo del axón hasta las siguientes neuronas. La velocidad de propagación depende de si el axón está o no recubierto por una sustancia aisladora llamada mielina. Para las neuronas no mielinizadas es de alrededor de 1 m/s, en tanto que para las neuronas recubiertas con mielina puede superar los 100 m/s. Una vez alcanzadas las terminaciones del axón, el potencial de acción se transmite a las otras neuronas a través de los contactos sinápticos. (La mielina reduce la capacidad eléctrica entre el axón y el medio exterior, disminuyendo así el tiempo necesario para cargar esa capacidad. Esto acelera la propagación de los pulsos).
Cuando se aplica un potencial constante en las dendritas de una neurona, el potencial de la neurona tiende a acercarse, con cierta constante de tiempo, al potencial aplicado. Si el potencial aplicado es suficientemente alto, en algún momento se supera el umbral y se produce el disparo, volviendo la neurona a su estado inicial. Después de completarse el período refractario, el ciclo vuelve a empezar, lo cual lleva a que se genere un tren de potenciales de acción. La frecuencia de este tren de pulsos aumenta al aumentar el potencial constante aplicado. Resulta, así, que la neurona se comporta en forma similar a un modulador de frecuencia, codificando las señales recibidas a través de la frecuencia de los potenciales de acción.
La señal que reciben las neuronas que inervan el oído interno proviene de las células sensoriales conectadas a la membrana basilar (células ciliadas internas). Cuando la membrana se encuentra en reposo (no es excitada por ningún sonido), estas células producen suficiente señal como para estimular una emisión espontánea de potenciales de acción con determinada frecuencia promedio. (El nivel umbral, así como la salida de las células sensoriales, están en realidad contaminados por una considerable cantidad de ruido, lo cual hace que la frecuencia de emisión fluctúe aleatoriamente alrededor de un valor medio). Cuando aparece un sonido, los movimientos hacia arriba y hacia abajo de la membrana basilar aumentan y disminuyen respectivamente el nivel de señal de las células sensoriales, de tal manera que en un caso la frecuencia aumenta y en el otro disminuye. Sin embargo, dado que el período refractario de las neuronas es del orden de 2 ó 3 ms, para frecuencias mayores de unos 300 Hz, esta variación de frecuencia no puede manifestarse directamente, sino que lo hace en forma estadística. El resultado es una tendencia a alcanzar una suerte de "sincronismo" entre la onda mecánica en la membrana basilar (y, por consiguiente, el sonido que la excita) y las emisiones de potenciales de acción. Este sincronismo se refiere a que los intervalos entre potenciales de acción tienden a ser múltiplos aproximados del período de la señal sonora. La corteza cerebral utiliza esta codificación para asignar una altura a un tono.
Es interesante destacar que las curvas de resonancia de la figura 9 son demasiado anchas para explicar la gran discriminación de frecuencias del oído humano. Este problema desconcertó durante años a los investigadores, hasta que se descubrió que la resonancia de la membrana basilar no obedece solamente a sus características elásticas pasivas, sino a un mecanismo de control en el cual intervienen las células ciliadas externas (figura 6) como elementos contráctiles. Estas células realimentan el sistema agudizando la resonancia.